
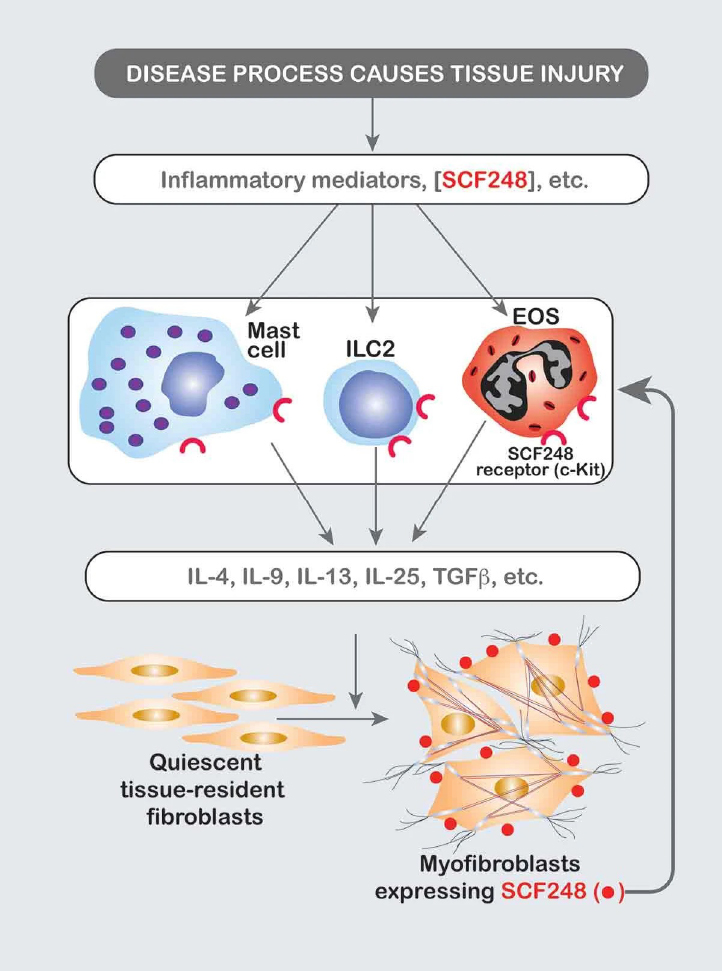
 Mast cells are multifunctional cells that can initiate or modulate various inflammatory processes. These cells contain many preformed mediators that, when released, may induce initial and necessary interactions between circulating leukocytes and the endothelium of postcapillary venules. The activation and degranulation of mast cell populations are responses that can be mediated by either antigen-specific, surface bound IgE or by stem cell factor (SCF). IgE-mediated mast cell activation induces immediate mast cell degranulation that constitutes the primary mechanism that drives the allergic responses and type I hypersensitivity. During this activation, mast cells release preformed and newly synthesized pro-inflammatory mediators including histamine, heparin, proteases, prostaglandin D2 , leukotriene C4 , and cytokines. In addition, murine mast cells challenged in an IgE-dependent manner or with other stimuli can produce multiple chemokines, including CCL3 (MIP1a), CCL4 (MIP-1b) and CCL2 (MCP-1) that can initiate and perpetuate an inflammatory cascade. Over the years we have identified the central role of mast cells in allergic inflammation and have further identified SCF as a primary target in peripheral tissue as being upregulated during inflammation and mediating chronic mast cell associated responses. Endogenous SCF occurs primarily in two forms, a 248 amino acid (AA) cleavable form (SCF248) and a 220 AA “noncleavable” form (SCF220) that differ by the presence or absence of exon 6 that encodes for protease cleavage site(s). Both isoforms of SCF are inserted into the plasma membrane, with the extracellular domain (ECD) of SCF248 more efficiently cleaved and shed from the surface of the cell during inflammation. While SCF-ECD is abundantly detected in circulation (~800 pg/mL in humans), circulating SCF-ECD is primarily monomeric that cannot cross-link and activate c-kit. SCF isoforms appear to play divergent roles in development, erythropoiesis, and myelopoiesis, where SCF220 may be required during development and erythropoiesis and SCF248 may be required for normal myelopoiesis and mast cell development and/or differentiation. We have furthermore identified that SCF248 is specifically upregulated in peripheral tissues during chronic inflammatory responses and promote the accumulation of mast cells, eosinophils and ILC2, all innate immune cells that express the SCF receptor, c-kit. The subsequent activation of fibroblasts and other non-immune cells by innate cell cytokines would drive disease severity as well as increase more SCF248 in a Feed-forward activation loop. The inhibition specifically of SCF248 with an antibody that recognizes the SCF248 isoform but not the SCF220 isoform, attenuates the development and severity of ongoing allergic and fibrotic disease models. Thus, we believe that targeting the SCF248 isoform will regulate chronic disease without altering the homeostatic functions of c-kit, such as erythropoiesis. (1-22)
Mast cells are multifunctional cells that can initiate or modulate various inflammatory processes. These cells contain many preformed mediators that, when released, may induce initial and necessary interactions between circulating leukocytes and the endothelium of postcapillary venules. The activation and degranulation of mast cell populations are responses that can be mediated by either antigen-specific, surface bound IgE or by stem cell factor (SCF). IgE-mediated mast cell activation induces immediate mast cell degranulation that constitutes the primary mechanism that drives the allergic responses and type I hypersensitivity. During this activation, mast cells release preformed and newly synthesized pro-inflammatory mediators including histamine, heparin, proteases, prostaglandin D2 , leukotriene C4 , and cytokines. In addition, murine mast cells challenged in an IgE-dependent manner or with other stimuli can produce multiple chemokines, including CCL3 (MIP1a), CCL4 (MIP-1b) and CCL2 (MCP-1) that can initiate and perpetuate an inflammatory cascade. Over the years we have identified the central role of mast cells in allergic inflammation and have further identified SCF as a primary target in peripheral tissue as being upregulated during inflammation and mediating chronic mast cell associated responses. Endogenous SCF occurs primarily in two forms, a 248 amino acid (AA) cleavable form (SCF248) and a 220 AA “noncleavable” form (SCF220) that differ by the presence or absence of exon 6 that encodes for protease cleavage site(s). Both isoforms of SCF are inserted into the plasma membrane, with the extracellular domain (ECD) of SCF248 more efficiently cleaved and shed from the surface of the cell during inflammation. While SCF-ECD is abundantly detected in circulation (~800 pg/mL in humans), circulating SCF-ECD is primarily monomeric that cannot cross-link and activate c-kit. SCF isoforms appear to play divergent roles in development, erythropoiesis, and myelopoiesis, where SCF220 may be required during development and erythropoiesis and SCF248 may be required for normal myelopoiesis and mast cell development and/or differentiation. We have furthermore identified that SCF248 is specifically upregulated in peripheral tissues during chronic inflammatory responses and promote the accumulation of mast cells, eosinophils and ILC2, all innate immune cells that express the SCF receptor, c-kit. The subsequent activation of fibroblasts and other non-immune cells by innate cell cytokines would drive disease severity as well as increase more SCF248 in a Feed-forward activation loop. The inhibition specifically of SCF248 with an antibody that recognizes the SCF248 isoform but not the SCF220 isoform, attenuates the development and severity of ongoing allergic and fibrotic disease models. Thus, we believe that targeting the SCF248 isoform will regulate chronic disease without altering the homeostatic functions of c-kit, such as erythropoiesis. (1-22)
 ON THE COVER
ON THE COVER
Breast team reviewing a patient's slide. (From left to right) Ghassan Allo, Fellow; Laura Walters, Clinical Lecturer; Celina Kleer, Professor. See Article |
newsletter
INSIDE PATHOLOGYAbout Our NewsletterInside Pathology is an newsletter published by the Chairman's Office to bring news and updates from inside the department's research and to become familiar with those leading it. It is our hope that those who read it will enjoy hearing about those new and familiar, and perhaps help in furthering our research. CONTENTS
|
 ON THE COVER
ON THE COVER
Autopsy Technician draws blood while working in the Wayne County morgue. See Article |
newsletter
INSIDE PATHOLOGYAbout Our NewsletterInside Pathology is an newsletter published by the Chairman's Office to bring news and updates from inside the department's research and to become familiar with those leading it. It is our hope that those who read it will enjoy hearing about those new and familiar, and perhaps help in furthering our research. CONTENTS
|
 ON THE COVER
ON THE COVER
Dr. Sriram Venneti, MD, PhD and Postdoctoral Fellow, Chan Chung, PhD investigate pediatric brain cancer. See Article |
newsletter
INSIDE PATHOLOGYAbout Our NewsletterInside Pathology is an newsletter published by the Chairman's Office to bring news and updates from inside the department's research and to become familiar with those leading it. It is our hope that those who read it will enjoy hearing about those new and familiar, and perhaps help in furthering our research. CONTENTS
|
 ON THE COVER
ON THE COVER
Director of the Neuropathology Fellowship, Dr. Sandra Camelo-Piragua serves on the Patient and Family Advisory Council. |
newsletter
INSIDE PATHOLOGYAbout Our NewsletterInside Pathology is an newsletter published by the Chairman's Office to bring news and updates from inside the department's research and to become familiar with those leading it. It is our hope that those who read it will enjoy hearing about those new and familiar, and perhaps help in furthering our research. CONTENTS
|
 ON THE COVER
ON THE COVER
Residents Ashley Bradt (left) and William Perry work at a multi-headed scope in our new facility. |
newsletter
INSIDE PATHOLOGYAbout Our NewsletterInside Pathology is an newsletter published by the Chairman's Office to bring news and updates from inside the department's research and to become familiar with those leading it. It is our hope that those who read it will enjoy hearing about those new and familiar, and perhaps help in furthering our research. CONTENTS
|
 ON THE COVER
ON THE COVER
Dr. Kristine Konopka (right) instructing residents while using a multi-headed microscope. |
newsletter
INSIDE PATHOLOGYAbout Our NewsletterInside Pathology is an newsletter published by the Chairman's Office to bring news and updates from inside the department's research and to become familiar with those leading it. It is our hope that those who read it will enjoy hearing about those new and familiar, and perhaps help in furthering our research. CONTENTS
|
 ON THE COVER
ON THE COVER
Patient specimens poised for COVID-19 PCR testing. |
newsletter
INSIDE PATHOLOGYAbout Our NewsletterInside Pathology is an newsletter published by the Chairman's Office to bring news and updates from inside the department's research and to become familiar with those leading it. It is our hope that those who read it will enjoy hearing about those new and familiar, and perhaps help in furthering our research. CONTENTS
|
 ON THE COVER
ON THE COVER
Dr. Pantanowitz demonstrates using machine learning in analyzing slides. |
newsletter
INSIDE PATHOLOGYAbout Our NewsletterInside Pathology is an newsletter published by the Chairman's Office to bring news and updates from inside the department's research and to become familiar with those leading it. It is our hope that those who read it will enjoy hearing about those new and familiar, and perhaps help in furthering our research. CONTENTS
|
 ON THE COVER
ON THE COVER
(Left to Right) Drs. Angela Wu, Laura Lamps, and Maria Westerhoff. |
newsletter
INSIDE PATHOLOGYAbout Our NewsletterInside Pathology is an newsletter published by the Chairman's Office to bring news and updates from inside the department's research and to become familiar with those leading it. It is our hope that those who read it will enjoy hearing about those new and familiar, and perhaps help in furthering our research. CONTENTS
|
 ON THE COVER
ON THE COVER
Illustration representing the various machines and processing used within our labs. |
newsletter
INSIDE PATHOLOGYAbout Our NewsletterInside Pathology is an newsletter published by the Chairman's Office to bring news and updates from inside the department's research and to become familiar with those leading it. It is our hope that those who read it will enjoy hearing about those new and familiar, and perhaps help in furthering our research. CONTENTS
|
 ON THE COVER
ON THE COVER
Rendering of the D. Dan and Betty Khn Health Care Pavilion. Credit: HOK |
newsletter
INSIDE PATHOLOGYAbout Our NewsletterInside Pathology is an newsletter published by the Chairman's Office to bring news and updates from inside the department's research and to become familiar with those leading it. It is our hope that those who read it will enjoy hearing about those new and familiar, and perhaps help in furthering our research. CONTENTS
|
MLabs, established in 1985, functions as a portal to provide pathologists, hospitals. and other reference laboratories access to the faculty, staff and laboratories of the University of Michigan Health System’s Department of Pathology. MLabs is a recognized leader for advanced molecular diagnostic testing, helpful consultants and exceptional customer service.